|
Migrace sladkovodních ryb
Úvod
Proč nacházíme živé organismy na
určitém místě? Proč se celá řada živočichů během svého života vydává na
mnohdy dlouhé nebezpečné cesty a jak to dokáží? Podobné otázky si musí
položit každý, kdo se stává cíleně nebo náhodně svědkem migrace velké
skupiny živočichů. Schopnost přemisťovat se z místa na místo patří mezi
základní vlastnosti živých organismů. A platí to i pro ty organismy, které
se nám zprvu mohou jevit jako zdánlivě nepohyblivé - například obří sekvoje
nebo přisedle žijící perlorodky říční - ale i ony v určité fázi života
vytvářejí formy, které jsou schopné přemístit se na několik desítek metrů
nebo i desítek či stovek kilometrů.
Migrace a
disperze
Migrace je hromadný pohyb
jedinců téhož druhu určitým směrem z jednoho místa na druhé. Migrace bývají
podmíněny geneticky a dále jsou určovány celou řadou vnitřních i vnějších
vlivů. Nejčastěji jsou migrace děleny dle jejich biologického účelu na
migrace potravní, rozmnožovací a migrace s cílem vyhledat vhodné útočiště.
Jedinci přitom mohou v průběhu svého života realizovat všechny typy migrací.
Mezi klasické migranty patří například tažné druhy ptáků, například rybák
dlouhoocasý (Sterna paradisea), který z Arktidy, kde vyvádí mladé táhne do
antarktidy a přitom urazí až 17 000 km. Nebo motýl monarcha (Danaus
plexippus), který migruje z jihu USA na sever a do Kanady, aby se zde
rozmnožil.
Disperze označuje vzdalování jedinců od jiných. Mezi disperzní pohyby patří
například vzdalování se potomků od svých rodičů, nebo semen od mateřských
rostlin a další.
Každý organismus či jeho populace, setrvávající po celý svůj život pouze na
jednom místě by byl dříve nebo později odsouzen k zániku. Život organismů
ovlivňuje celá řada faktorů prostředí, které se neustále mění. Přežití
stávajících i budoucích jedinců a populací pak na neustálém vyhledávání
optimální podmínek k životu. Migrace tedy sehrávají u živých organismů
klíčovou roli.
Dělení migrací
ryb
Již od dětství každý z nás
slýchal o dlouhých migracích lososů, kteří se vydávají na dlouhou cestu z
moří do velkých řek a dále do menších přítoků hluboko do vnitrozemí, kde se
třou a následně (u některých druhů) pak celé tisíce dospělých jedinců v
průběhu několika málo dní hyne. Stejně tak jsou všeobecné známy migrace
úhořů opačným směrem z řek do oceánu a pak do Sargasového moře, kde ve
velkých hloubkách dochází k jejich rozmnožování. Délky těchto migrací
dosahují tisíců kilometrů a jsou spektakulárním přírodním divadlem. Skoro by
se zdálo, že jsou zbytečným mrháním přírody, ale při hlubším poznání se
jedná o velmi důmyslný mechanismus přežití jednou druhu uprostřed
nejrůznějších vlivů prostředí a konkurence dalších organismů.
Migrace můžeme rozdělit dle
období jejich konání na
1. pravidelné – zpravidla sezónní (např. v období tření) nebo denní (cirkadiánní
opakující ze dne na den například v závislosti na pohybu planktonu v
jezerech)
2. nepravidelné – vyvolané změnou faktorů prostředí (například nepříznivou
změnou chemismu vody)
Dle aktivity migrujících jedinců můžeme rozlišit migrace:
1. aktivní
2. pasivní
Aktivní migrace vyžadují
svalovou aktivitu jedinců. Tyto migrace provází výdej energie. Výhodou
aktivních migrací je jejich směrování proti působení fyzikálních sil.
Odehrávají se proti proudu i za poměrně vysokých rychlostí proudění. Ryby
mohou při aktivních migracích překonávat i příčné překážky. Například pstruh
obecný je při svých migracích v době tření překonat skokem překážku až do
výše 1,15 m. Aktivní migrace umožňují rybám vyhledat vhodné podmínky pro
nakladení jiker a vývoj raných vývojových stádií. Pasivní migrace se
odehrávají za působení fyzikálních sil (proudění vody apod.) a pro pasivně
migrujícího jedince nepředstavuje tento způsob zvláštní energetické nároky.
Pasivní migrace jsou typické zejména pro juvenilní stádia ryb, která bývají
z míst líhnutí z jiker unášena proudem a usazují se ve vyhovujících
podmínkách. Tím je také zajištěna jeho rovnoměrná distribuce v prostředí
(tzv. kompenzační migrace). Proudění vody je typické nejen v tekoucích
vodách, ale i ve vodách stojatých a mořích. Mořských proudů využívají k
migraci například plůdek sledě obecného (Clupea harengus). Unášen mořským
proudem může urazit vzdálenost až 1.000 km.
Podle směru pohybu ryb v
prostoru rozeznáváme migrace:
1. horizontální
2. vertikální
V tekoucích vodách jsou
lineárním ekosystémem s charakteristicky uspořádaným prostředím a proto také
migrace ryb se v tekoucích vodách odehrávají v horizontální rovině po
proudu, protiproudu a nebo laterálně v době záplav, kdy se rybám
zpřístupňují rozsáhlé záplavové území mimo hlavní koryto toku.
Ve stojatých vodách nebo velkých řekách s větší hloubkou se pak setkáváme s
vertikálními migracemi. Takové migrace mohou být motivovány potravou
(například planktonu, který se shromažďuje během dne v prosvícených a
prohřátých svrchních vrstvách a v noci klesá do větších hloubek), únikem
před predátorem, přezimování ve větších hloubkách, nebo vyhledávání vrstev
vody s optimální koncentrací kyslíku.
Dlouhé tahy ryb se odehrávají nejen v prostředí sladkých vod nebo moří, ale
také mezi nimi. Podle prostředí tak rozeznáváme migrace:
1. Oceanodromní
Migrace odehrávající se v mořském prostředí. Např. platýs velký (Pleuronectes
platesa).
2. Potamodromní
Migrace odehrávající se pouze ve sladkých vodách. Například pstruh obecný (Salmo
trutta m. fario) nebo mihule potoční (Lampetra planeri).
3. Diadromní
Migrace probíhající mezi sladkými vodami a mořem. Například losos obecný (Salmo
salar).
Diadromní migrace popisují migraci ryb mezi prostředím sladkých vod a mořem
většinou za účelem rozmnožování. Diadromní ryby však mohou podnikat také
další migrace, třeba za účelem vyhledávání potravy. Například mladí jeseteři
druhu Acipenser oxyrhinchus náležející ke skupině anadromních ryb podniká
letní migrace do brakických vod nebo přímo do řek o mnoho let dříve než
dosáhne pohlavní zralosti.
Typy diadromních ryb:
1. Anadromní
Ryby žijící většinu života ve slané vodě, kde se odehrává větší část jejich
životního cyklu. Do sladkých vod vstupují nejčastěji už ve stádiu pohlavní
dospělosti a pouze za účelem reprodukce (tření). Ve sladkých vodách už
prakticky nevyhledávají potravu a nepřirůstají (losos obecný).
2. Katadromní
Jsou opakem anadromních ryb. Většina ontogenetického vývoje probíhá ve
sladké vodě, ve slané vodě dochází pouze k rozmnožování (úhoř říční).
3. Amfidromní
Ryby, u kterých dochází k migraci do slané vody bezprostředně po vylíhnutí.
Juvenilní stádia pak migrují zpět do sladké vody, kde dospívají a probíhá
zde také jejich tření.
Rozsáhlý výzkum oceanodromních a diadromních migrací byl podnícen zejména
zájmy komerčního rybářství, které výsledky výzkumu aplikuje. Tento výzkum
zaměřený zejména na tahy ryb jako jsou lososi přispěl bezesporu k bližším
poznání ekologie, fyziologie a evoluce ryb, nicméně o to více byl opomíjen
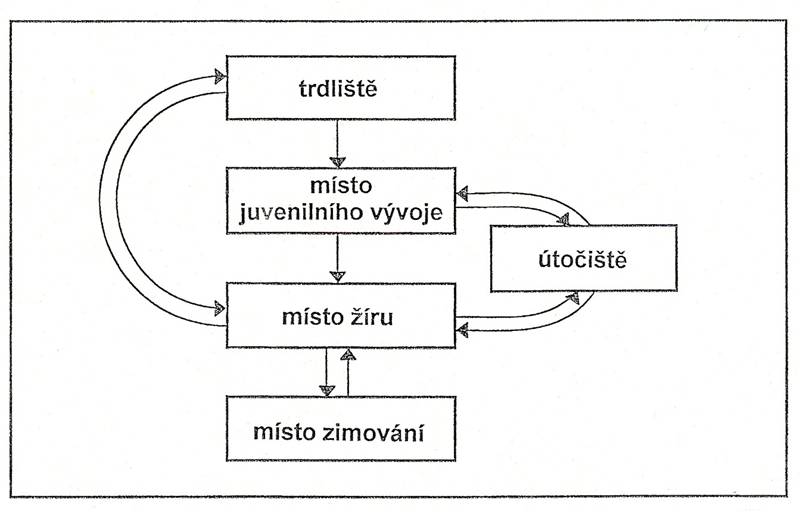
výzkum migrací potamodromních ryb.
Migrace odehrávající se v řekách či jezerech na vzdálenost desítek či stovek
metrů mohou být pro jedince stejně životně důležité, jako migrace konané na
vzdálenosti tisíců kilometrů. Pro některé druhy ryb může jeden habitat
naplnit veškeré požadavky, které u něj v průběhu životního cyklu vznikají.
Mnohé druhy evropských rheofilních druhů kaprovitých ryb jako například
parma obecná (Barbus barbus) využívá stejných lokalit ke tření, vyhledávání
a příjmu potravy, ale v nepříznivém období v průběhu zimy se vydává hledat
vhodnější podmínky. Jiné druhy zase svá zimoviště mají i jako vhodná místa
pro tření a za potravou se pak vydávají na lokality vzdálené. Naproti tomu
vranka obecná (Cottus gobio), nebo mřenka mramorovaná (Neomacheilus
barbatulus), které patří k méně zdatným plavcům a žijí benticky (při dně)
realizují tření, příjem potravy a zimování na jednom místě.
Přizpůsobení ryb k migracím
Třída ryb zahrnuje tvarově
poměrně různorodou skupinu živočichů, jejíž zástupci obývají takřka výhradně
vodní prostředí, kterému jsou dokonale přizpůsobeni. Právě díky širokému
spektru morfologických a fyziologických adaptací patří ryby mezi
nejpohyblivější zástupce vodní fauny.
Tvar těla
Tvar těla ryb je přizpůsoben
životu ve vodě. Tělo je zpravidla vřetenovitě protáhlé, ze stran pak
víceméně zploštělé a má odpovídající hydrodynamické vlastnosti. Rychle
plovoucí druhy ryb mají tělo protáhlé, na příčném průřezu oválné a zpravidla
bez tělních přívěsků (například štika obecná), které by zvyšovaly tření
rybího těla. Naopak tělo ryb pomalu plovoucích je kratší a vysoké (cejn
velký).
Pohybová soustava
Svalstvo ryb, spoluutvářející
pohybovou soustavu, je tvořeno třemi různými typy svalových vláken. Jsou to
„červená“ vlákna s vysokým obsahem krevního barviva schopného vázat kyslík a
vlákna „bílá“ (tzv. glykolytická) a růžová vlákna. Červená svalová vlákna
jsou pomalá (pomalá reakce na podnět), pracují v aerobním režimu a jsou tedy
velmi dobře prokrvována soustavou kapilár. Jejich hlavní vlastností je
vytrvalost. Naopak bílá vlákna reagují na podnět velmi rychle, zdrojem
energie pro jejich činnost je glukóza vznikající anaerobním štěpením
glykogenu (glykolýza). Bílá vlákna pracují s vysokým výkonem, ale jen velmi
krátce (10 – 15 s). Červená svalová vlákna tvoří zpravidla menší část
svaloviny (3-5% u štik), ale u vytrvalých plavců, jako jsou lososi a některé
druhy rheofilních (proudomilných) ryb to může být 15 – 20 %. Většina ryb
však mívá ve svalstvu největší podíl růžových svalových vláken, která
kombinují vlastnosti obou předešlých typů.
Spolu s ploutvemi tvoří svalstvo efektivní pohybový aparát zajišťující
stabilitu rybího těla ve vodním prostředí a také dostatečně rychlý pohyb.
Mezi nejrychlejší druhy ryb patří tuňák obecný (Thunnus thynnus) plovoucí
rychlostí až 6 m/s (cca 22 km/h) a losos obecný (Salmo salar), který se
dokáže krátkou dobu pohybovat rychlostí 5 m/s (18 km/h). Pstruh obecný (Salmo
trutta m. fario) dokáže vyvinout maximální rychlost okolo 4 m/s (cca 14
km/h). Průměrná rychlost i u zdatných plavců však bývá nižší (tažné formy
pstruha dosahují rychlostí okolo 10 km/h).
Tělní pokryv
Kromě tvaru těla a výkonného
pohybového aparátu usnadňuje pohyb ryb ve vodním prostředí také tělní
pokryv. Pokožka ryb je bohatě prostoupena žlázami produkujícími sliz. Sliz
má ochrannou funkci a také významným způsobem snižuje tření těla při pohybu
ve vodě. Tělo ryb může být lysé, ale velice často jej kryjí šupiny, které
jsou v pokožce zapuštěny a ovlivňují hydrodynamické vlastnosti rybího těla.
Rozlišujeme šupiny ganoidní, cykloidní a ktenoidní, které se liší svou
stavbou. Ktenoidní šupiny a cykloidní šupiny jsou ve tvaru jemných
zaokrouhlených, střechovitě se překrývajících destiček. Pokud je jejich
okraj hladký, hovoříme o cykloidních šupinách, pokud je zadní okraj posázen
jemnými zoubky, jedná se o šupiny ktenoidní. Ktenoidní šupiny ve srovnání s
cykloidními mají lepší hydrodynamické vlastnosti. Jejich povrchové, v
podélném směru uspořádané jemné zvrásnění, umožňuje lepší obtok vody a tedy
menší tření. Ktenoidní šupiny jsou typické pro rychle se pohybující ryby
např. candáta obecného (Styzostedion lucioperca), cykloidní šupiny pak pro
druhy pomalé např. kapra obecného (Cyprinus carpio).
Fyziologie
Vzdálenost migrací je dána nejen
pohybovou vybaveností migrujícího jedince, ale také schopností pokrýt
energetické nároky migrací. Lososi po dobu migrace nepřijímají potravu
vůbec. Plavou až téměř do úplného vyčerpaní své energetické rezervy a to je
překvapivě tajemstvím úspěšného dosažení jejich cíle. Ryby, které naopak v
průběhu migrace doplňují energii příjmem potravy totiž musí věnovat určitý
čas jejím vyhledáváním a příjmem, zejména v neznámém prostředí. Zpracování
potravy navíc představuje energetické výdaje, které nemohou být investovány
do plavání. Tyto faktory mohou zapříčinit nestihnutí optimálního doby a
podmínek pro tření.
Jako zdroj energie v době migrace mohou ryby využít cukry, bílkoviny a tuky,
přičemž poslední zmíněné jsou pro ně nejbohatším zdrojem energie. Ryby
podnikající dlouhé migrace se dokáží energeticky bohatými lipidy předzásobit
jejich ukládáním v těle.
Některé mořské druhy platýzů dokáží během migrace své energetické zásoby
prakticky úplně vyčerpat. Jejich svalstvo pak obsahuje až 95% vody, 3%
bílkovin a 0,05% tuku. S tak napjatou energetickou bilancí se u
potamodromních ryb nesetkáváme a to také proto, že by úplným vyčerpáním
mohla být ohrožena úspěšnost tření. Přesto jsou energetické výdaje třeba u
lososů značné a dosahují až 82 %. Ztráty hmotnosti se pak u lososoa obecného
pohybují po vytření v rozmezí 30 – 40 %.
U diadromních ryb vyvstává otázka, jak dokáží tolerovat tak velké rozdíly v
salinitě vodního prostředí při migracích z řek do moří a naopak. Není
automatické, že diadromní ryby jsou euryhalinními živočichy, tedy s velkou
tolerancí ke kolísání koncentrace rozpuštěných solí. Některé druhy určitě,
ale velká část z nich ne. Mnohem více se tedy jedná o fyziologickou
adaptaci. Například u lososa fyziologická adaptace spočívá ve změně činnosti
osmoregulačního mechanismu v průběhu ontogenetického vývoje z hyprosmotické
osmoregulace na hypoosmotickou v průběhu smoltifikace (smoltizace = změny ve
zbarvení, chování a metabolismu u juvenilních lososů) . Smoltifikace se
ovšem odehrává ještě v řekách, tedy dříve než ryby odplují do moře a přináší
rybám obtíže v podobě iontové disbalance snížení pohyblivosti a dalších.
Naopak jim toto přizpůsobení umožňuje rychleji proniknou brakickou zónu a
zachovat homeostázu při průniku do slané vody.
Orientace
Migrace v říčních ekosystémech
je omezena lineárním charakterem prostředí spolu s dalšími faktory -
rychlostí proudění, hloubkou vody, šířkou koryta. Orientace v prostředí pak
probíhá dle orientačních bodů, proudů, polarizovaného světla,
geomagnetického pole, chemických vlastností vody a další.
Orientační body v korytě toku nebo na březích jsou pro ryby při orientaci
bezesporu nepostradatelné v domovských okrscích. Například střevle potoční (Phoxinus
phoxinus) se ve svém prostředí velmi dobře vizuálně orientuje. Méně
významnou roli hrají orientační body při dlouhotrvajících tazích na delší
vzdálenost, nebo na otevřených vodách (jezera, velké řeky apod.).
Významným orientačním bodem je slunce, respektive jeho pozice. Nicméně v
době vyšší oblačnosti, kdy sluneční kotouč není vidět, musí být ryby schopny
využívat také jiné způsoby orientace. Slunce je zdrojem světla a
polarizované světlo slouží rovněž k účinné orientaci. Polarizované světlo
přitom vzniká v době úsvitu či soumraku, kdy je více než 60% světla
dopadajícího do vody může být polarizováno. Orientace podle polarizovaného
světla byla prokázána u mnoha druhů ryb.
Jedním z hlavních faktorů orientace v řekách, ale i v jezerech rozhodující
pro migraci ryb, je proudění. Většina ryb, zejména rheofilních je vybavena
vrozenou reotaxí, neboli chováním, kdy se staví hlavou proti proudu.
Intenzita proudění má vliv například na vstup lososů do ústí řek, je
významným orientačním faktorem pro ryby ve větších řekách.
Při pohybu vody napříč magnetickým polem Země může být indukováno elektrické
pole a přestože je jen velmi slabé může být zachyceno migrujícími rybami.
Právě tento jev má snad největší význam pro migraci v mořském prostředí, kde
další způsoby orientace mohou selhávat. Receptory indukovaných proudů byly
izolovány pletiv v oblasti nosní dutiny několika ryb. Podle magnetického
pole se dokáže orientovat například pstruh duhový (Oncorhynchus mykiss).
Magnetoreceptorové buňky pstruha duhového lokalizované v trojklaném nervu
obsahují krystaly magnetitu, který svým pohybem v magnetickém poli vyvolává
nervové vzruchy.
Čich hraje velmi důležitou roli a v devatenáctém století bylo zjištěno, že
je základním mechanismem u lososů, který řídí jejich návrat do domovských
řek. Ryby mohou být citlivé buď na kombinaci látek, které detekují jejich
prostředí, nebo si vštěpují pouze jednu specifickou látku. Citlivost ryb na
některé látky rozpuštěné ve vodě je obrovská. Přirozeně se vyskytující látky
jako aminokyseliny, žlučové kyseliny a soli a celá řada hormonů jsou
čichovými stimulátory a v některých případech mohou být detekovány v
koncentracích 10-16 M, což znamená, že zachycení několika málo molekul
těchto substancí vyvolává odezvu. U lososů je tato čichová paměť nastavena v
juvenilním stádiu a to ve velmi krátkém časovém úseku. Pokusy bylo dokázáno,
že toto nastavení lze uměle ovlivnit.
Homing – samonavádění
Je chování, kdy se ryby
spontánně vrací do domovských okrsků, odpočinková či třecí místa. S tímto
chováním jsou spojeny specifické mechanismy orientace a navádění. Jedná se
tedy o jakousi prostorovou paměť.
Toto chování bylo dobře zdokumentováno zejména u lososů, ale prokázáno také
u celé řady dalších druhů jako střevle potoční (Phoxinus phoxinus), kapra
obecného (Cyprinus carpio), parmy říční (Barbus barbus). Úspěšnost tohoto
chování může být velmi vysoká. U lososů překračuje 95 %, u dalších
lososovitých ryb pstruha duhového (Oncorhynchus mykiss) 94 %, pstruha
obecného 99,5% au sivena amerického (Salvenius fontinalis) dokonce až 100%.
Obecně platí, že přesnost je u lososovitých vyšší než u ostatních druhů.
Homming se může objevovat i u ryb podnikajících potravní migrace (parma
obecná, mník jednovousý).
U ryb můžeme hovořit o krátkodobé, střednědobé a dlouhodobé paměti.
Krátkodobá a střednědobá paměť jsou vyžívány k orientaci v domovském okrsku,
případně při krátkých migracích za potravou. Dlouhodobá paměť vzniká u ryb v
určité životní fázi. Například, jak už bylo zmíněno, čichová paměť lososů se
vštěpuje v krátkém období v průběhu juvenilní fáze. Pravděpodobně všechny
ryby jsou schopny vytvořit si jakýsi druh dlouhodobé paměti o pozici a stavu
domovského prostředí a to většinou ve velmi rané fázi vývoje, kdy začíná
fungovat nervová soustava. Dalším vysvětlením může být také zvýšená hladina
thyreotropního hormonu u juvenilních stádií ryb, která umožňuje jak jednak
vysokou citlivost organismu (mozku) na podněty, ale také vysokou schopnost
pamatovat si. Tímto se zřejmě zakládá dlouhodobá paměť.
Faktory ovlivňující migrace
Migrace jsou u ryb ovlivňovány celou řadou endogenních a exogenních faktorů.
Genetické faktory
Migrační chování má genetický
základ a jeho projev je velmi silně ovlivňován faktory prostředí a
ontogenetickým vývojem. U diadromních druhů lososovitých ryby je migrační
chování silně vyvinuto, ale jeho projevy se liší mezi populacemi v různých
řekách, což je ovlivněno například množstvím a dostupností potravy. U
driftujících juvenilních stádií některých tropických druhů ryb probíhají
migrace zejména v noci nebo v době zákalu vody, aby tak snížily potenciální
riziko ulovení predátorem.
Metabolismus
Potrava je pro ryby stejně jako
pro ostatní živočichy zdrojem energie pro uskutečnění životních potřeb. Z
fyziologického hlediska jsou potravní migrace vyvolány drážděním trávicího
traktu (zejména žaludku a střeva) přítomností potravy a současně
metabolickou bilancí, která je kontrolována složitým regulačním systémem,
který hlídá hodnotu energeticky bohatých látek v organismu.
Potravní migrace se odehrávají horizontálně i vertikálně. Vertikální migrace
mohou mít přitom různé důvody. Může to být sledování kořisti – pelagicky
žijící bezobratlé (plankton) – při jejich diurnálních migracích, nebo snaha
vyhnout se predátorům. Některé teorie hovoří také o tom, že účelem těchto
migrací mohou být energetické úspory. Ryby totiž patří mezi poikilotermní
organismy a teplota jejich těla se blíží teplotě prostředí. V teplejších
povrchových vrstvách proto metabolismus probíhá rychleji a energie získaná
potravou je tak spotřebována rychleji. Naopak v hlubších chladných vrstvách
dojde k výraznému zpomalení životních procesů a ryby tak mohou třeba přečkat
nepříznivé období provázené nedostatkem potravy.
Migrace obecně jsou pro organismus energeticky velmi náročné. Jedinec si
tedy musí před nástupem dlouhé migrace vytvořit dostatečnou energetickou
zásobu. Délka migrace je pak přímo úměrná energetickým možnostem organismu.
Celá řada druhů se tedy nevydává na cestu dříve, než si takovou rezervu
nevytvoří.
Predace
Mnoho ryb migrujících za účelem
nalezení nových vhodných útočišť tak činí ve snaze vyhnout se riziku ulovení
predátorem. Tento faktor náleží spíše mezi vnější, ale u jedince se může
projevit až na základě získaných zkušeností a učení. Jejich manifestace je
pak faktorem vnitřním. Signály k započetí takového chování jsou různé.
Některé druhy ryb z řádu maloostných vylučují do prostředí zvláštní látku,
která se uvolňuje z poškozených tkání v okamžiku napadení predátorem. Tyto
látky pak mohou zachytit ostatní ryby a reagovat únikem. K projevům migrací
z důvodu úniku před predátorem však může být jeho pouhá přítomnost
registrovaná zrakem a jinými receptory. Predačním tlakem (např. při
přemnožení predátora) tak může postupně dojít ke změně struktury
společenstva ve prospěch ryb, které nejsou tak snadnou kořistí.
Světlo
Biologické cykly ryb jsou
ovlivňovány střídáním světelné a tmavé fáze dne. Reakce mohou být přímé nebo
nepřímé. Například nepřímou reakcí jsou vertikální migrace ryb v jezerech za
kořistí – planktonem – , který migruje ve dne k hladině a v noci do hloubky.
Jelec proudník (Leuciscus leuciscus) se pravidelně se soumrakem vrací do
proudných úseků, kde stráví noc a před úsvitem se vrací na mělčinu. Světlo
pro ryby znamená také vyšší riziko ulovení predátory, kteří využívají k
orientaci zrak, proto jsou některé druhy ryb aktivní zejména v noci. Je tedy
zřejmé, že světlo má v přirozeném prostředí přímý vliv na migraci ryb a je
tedy zřejmé, že faktory snižující intenzitu světla ve vodě ovlivňují také
migraci. Jedná se například o zákal – turbiditu. Například lososi při svých
tazích vstupují do řek téměř výhradně v noci. Pokud je však voda v řece
zakalená, což bývá spojeno také s vyšší rychlostí proudu (například při
zvýšených průtocích) migruje také během dne, neboť tyto podmínky jsou pro
něj optimální. Také larvální a juvenilní stádia tropických druhů ryb
driftující po proudu obvykle v noci, vstupují do vodního sloupce také přes
den, je-li voda zkalena.
Teplota
Ryby jsou poikilotermní
živočichové a proto jsou aktivní zejména při vyšších teplotách. Teplota vody
je tedy faktorem, který určuje také periodicitu dlouhých migrací ryb a
tření. Už malé výkyvy v teplotě vody mohou vyvolat změny chování ryb v
průběhu tahu či tření. Prahová hodnota teploty může být u některých druhů
velice přesně daná. Například mihule potoční (Lampetra planeri) se v
norských řekách vydává na protiproudové třecí migrace při dosažení teploty
vody 7,5 °C. Teplotní interval se v našich podmínkách však obecně bývá u
tohoto druhu širší (8 – 15 °C). Celá řada studií larev úhoře říčního
prokázaly, že migrace do řek začíná při teplotě 6-8 °C. V Labi pak byla
prokázána zvýšená aktivita úhořů při tahu v okamžiku poklesu teploty vody
pod 10 °C.
Hydrologie a meteorologie
Mnoho druhů ryb mírného pásma,
zejména lososovitých ryb, začíná migraci v období zvýšených průtoků. Zvýšené
průtoky v jarním obdob způsobené vyššími srážkami nebo táním sněhu mohou
zapříčinit záplavy, které stimulují larvy, juvenilní stádia ryba a pohlavně
dospělé jedince k laterálním migracím do záplavových oblastí. Ve sníženinách
a příkopech, kde se voda po záplavách udrží nejdéle, pak dochází ke tření
ryb. Takové chování je typické například pro štiku obecnou (Esox lucius). U
některých kaprovitých ryb tropických oblastí je tření načasováno přímo na
dobu průtrže mračen a bouřek. Pokud dojde k jejich zpoždění, nebo nepřijdou,
může proces tření úplně selhat. Při překročení určité výše průtoku však
mohou migrace naopak ustávat, například u lososovitých ryb nebo u mihule
potoční, která dokola překonat rychlost proudu pouze do 2 m/s. Důvodem je
pravděpodobně vysoká energetická náročnost pohybu proti silnému proudu.
Sezónní fluktuace průtoků je v rámci říčních ekosystémů velmi významným
dynamickým faktorem, který zajišťuje vyváženou distribuci rybích populací.
Kvalita vody
Kvalita vody je určována buď
přirozenými vlastnostmi prostředí nebo antropogenními aktivitami. Například
když dojde při záplavách k rozlivům do lužního lesa a ve vodě se začne
rozkládat opadlé listí, může dojít k významnému snížení či úplnému vyčerpání
kyslíku. Člověk kvalitu vody zhoršuje plošnými či bodových zdrojů znečištění
různého typu. Migrace ryb pak reflektují změny v kvalitě vody a jejich cílem
bývá zpravidla únik před dlouhodobě či krátce nepříznivými podmínkami.
Změnou v kvalitě vody byly podníceny také migrace kaprovitých ryb mezi
hlavním korytem a slepými rameny ve Vltavě. Ve dne se ryby shromažďovaly ve
slepých ramenech, kde v důsledku fotosyntézy řas a rostlin bylo dostatek
kyslíku. V noci se ryby naopak vracely do hlavního koryta, protože v
důsledku respirace (dýchání) vegetace došlo ve slepých ramenech k postupnému
vyčerpání kyslíku. Podobné chování bylo zaznamenáno také u hlavatky
podunajské (Hucho hucho) na Slovensku.
Potravní zdroje
Jak již bylo dříve zmíněno,
migrace za potravou jsou vyvolávány bilancí metabolismu. Jejich cílem je
tedy zajistit zdroj látek, které umožní realizaci životních pochodů v
organismu. Migrace za potravou je určována zejména její dostupností. Ryby se
vydávají za potravou až v okamžiku, kdy je jí v domovském okrsku nedostatek.
U juvenilních stádií hlavatky podunajské byly zjištěny poproudové migrace a
jejich aktivní vyhledávání míst s koncentrací plůdku jiných druhů ryb,
kterými se živili.
Biologický účel migrací
Migrace jsou již tradičně
rozdělovány dle tří hlavních účelů na migrace potravní, migrace,
rozmnožovací a migrace na útočiště. Je však zřejmé, že toto přehledné dělení
neodpovídá složité provázanosti jednotlivých podnětů, stimulujících ryby k
migracím a mnohdy nezřetelné hranice mezi jednotlivými typy. To je potřeba
mít při popisu jednotlivých typů migrací na paměti.
Potravní migrace
Potravní migrace jsou velmi
různorodé a jejich projevy mohou být u různých druhů zcela odlišné a
probíhající v různých obdobích roku. Potravní migrace jsou také spojeny s
ontogenetickým vývojem ryb. Ryby migrují za potravou zpravidla v obdobích
tělesného růstu a rozvoje a je tedy pravděpodobné, že potravní migrace více
korespondují s těmito důvody než s pouhou snahou maximalizovat příjem
potravy. Přesto však lze pozorovat, že ryby ve svých migracích sledují také
produktivitu jednotlivých ekosystému, například rozdíly mezi marinním a
sladkovodním ekosystémem. Také potamodromní migrace sledují lokality s
různou úrovní produkcí. Například laterální migrace do záplavových území
mají z hlediska uvolnění velkých zásob živin pro ryby velký význam.
U některých anadromních a většiny amfidromních ryb je běžné, že jejich jikry
jsou splaveny z řek do moře, kde rostou, vyvíjejí a dospívají. U
sladkovodních ryb jsou zpravidla ryby při kladení připevněny k podklady a po
proudu migruje až vylíhlý plůdek. U lososů je ranný plůdek již dobře vybaven
k pohybu. Plůdek některých amerických druhů lososů migruje bezprostředně po
vylíhnutí. Losos obecný a pstruh obecný začíná poproudovou migraci až po
transformaci ve smolty. Pasivní migrace plůdku pstruha obecného byla
pozorováa jen za nepříznivých hydrologických podmínek (vyšší průtoky).
Protiproudová migrace juvenilních ryb je charakteristická zejména pro
katadromní (úhoř) a některé amfidromní druhy ryb.
Dospělci podnikají potravní migrace, které jsou často typické pro určité
období roku, například na jaře nebo v létě. Mohou být také spojeny s
migracemi za účelem tření. Například některé lososovité ryby po vytření
migrují na lokality s dostatečnou potravní nabídkou, aby doplnily energii
vydanou při náročném tahu. Studie u mnoha běžných druhů kaprovitých ryb jako
cejn velký, parma říční, jelec tloušť a dalších prokázaly, že v letním
období se délka migrací pohybuje okolo 3 km a jen výjimečně jsou delší.
Stejně tak štika obecná podniká taky za potravou jen velmi zřídka a jejich
vzdálenost dosahuje nejčastěji maximálně 2 km. Je však zajímavé, že jedinci
podnikající delší migrace byla zjištěna větší délka i hmotnost těla, což
bylo způsobeno pravděpodobně větší zabezpečeností potravou. Kvalita potravy
může být pro ryby stimulem k migraci. Část populace plotice obecné v
některých nádržích se může po ukončení třecích migrací v přítocích s
odstupem jednoho nebo více měsíců migrovat opětovně do přítoků z důvodu
kvalitní potravní nabídky, která je převládající živočišnou složkou
atraktivnější než převážně rostlinná strava v jezeře.
Migrace za útočištěm
V případě změny podmínek
prostředí, které jsou pro ryby nevýhodné, je nezbytné, aby se takové změně
adaptovaly, nebo se pokusili nalézt nové útočiště s vhodnějšími podmínkami.
Podobné chování bylo velmi důležité také v období střídání geologických
období a pomáhalo přežití rybích populací při změně klimatu například v době
ledové.
Hlavatka podunajská (Hucho hucho) reaguje na krátkodobé nepříznivé zhoršení
prostředí poproudovými nebo protiproudovými migracemi do přítoků. Jakmile
dojde ke zlepšení podmínek vrací se na svá stanoviště.
Snaha vyhnout se nepříznivým podmínkám v zimním období je typická pro úhoře
říčního. V řekách, brakických vodách nebo přílivových zónách úhoři migrují
do slepých ramen, kanálů a příkopů, kde je dostatečně hluboká, aby úhoře
ochránila před nepříznivými účinky zimy. Takové migrace mohou dosahovat 2 –
5 km.
V mírném pásmu jsou typické poproudové migrace ryb z plyších a proudných
úseků do klidnějších a hlubších partií roku, kde přezimují. Pokud je
morfologie říčního koryta dostatečně pestrá, nemusí k takovým migracím vůbec
docházet a ryby mohou přezimovat na identických lokalitách, které obývají v
létě. Takové chování je ryby výhodné, protože migrace vždy představují
riziko snížení tělesné kondice a onemocnění, případně riziko ulovení
predátorem. Poproudové migrace s účelem přezimování jsou typické třeba pro
cejna velkého nebo candáta obecného. Do nádrží se před zimou se mohou
přesouvat také juvenilní stádia lososovitých ryb, které vyhledávají
štěrkovité nebo kamenité dno.
Nepříznivé podmínky pro ryby v tocích mohou nastávat také v období sucha,
kdy klesají průtoky k dlouhodobým minimům. Tehdy se ryby shromažďují v
přítocích nebo tůních. Některá druhy mohou v situacích, kdy dochází k
vyschnutí koryta migrovat (zavrtávat se) do dna (hyporeické zóny) jako
například benticky žijící vranka obecná (Cottus gobio). V některých
případech byla zjištěna v hloubce až 1 m.
Třecí migrace
Cílem třecích migrací je oddělit
juvenilní a adultní stádia z důvodu jejich rozdílných životních nároků a
také z důvodu rovnoměrné distribuce vyvíjejících se jedinců při jejich
poproudových migracích. Migrace za třením jsou ve srovnáni s předešle
zmiňovanými typy velmi jasně zřetelné, protože probíhají u jednotlivých
druhů většinou v krátkém jasně vymezeném období roku a jsou provázeny
nahlučením jedinců obou pohlaví. Pro celou řadu litofilních druhů mezi které
patří například kruhoústí, lososovité, okounovité a kaprovité ryby, které si
vytloukají hnízda pro kladení jiker ve štěrkovém substrát patří
protiproudová migrace za třením mezi charakteristické vzorce chování.
Migrace přitom mohou být velmi krátké jako v případě druhů, přezimujících v
nádržích, nebo velmi dlouhé jako v případě amerického lososa Oncorhynchus
nerca. Štika obecná v době tření podniká migrace dosahující délky až 20 km.
Migrace pstruha obecného jsou spíše krátké a jen zřídla překračují 1 km. V
táhnoucích hejnech jsou pozorovány nejdříve samci, samice až později. Z
kaprovitých ryb patří mezi nejaktivnější migranty v době tření ostroretka
stěhovavá. I u tohoto druhu se však velká část populace nevzdaluje svému
domovskému okrsku (dle některých údajů až 77 % populace), zcela ojediněle
byly zaznamenány tahové vzdálenosti v řádu stovek kilometrů.
Jedním z cílů migrací je také zajistit rovnoměrnou distribuci jedinců v
rámci vodního toku. Proto je jedním z určujících faktorů pro délku migrací
také typ jejich jiker případně způsob života jejich juvenilních stádií. U
druhu se vzplývavými jikrami jsou protiproudové migrace zpravidla velmi
dlouhé a to z toho důvodu, aby byla zajištěna dostatečná vzdálenost pro
jejich vývoj a usazení v optimálních podmínkách v průběhu jejich driftu po
proudu. Takové migrace jsou typické pro některé druhy toků tropické Afriky a
Jižní Ameriky. Jiné druhy se vzplývavými jikrami zase migrují do slepých a
mrtvých ramen nebo záplavových území.
Migrace ryb a člověk
Ryby jsou a v minulosti byly pro
člověka významným zdrojem potravy. Řeky vždy znamenaly strategické místo pro
zakládání lidských sídlišť z důvodu jejich mnohostranného využití.
Atraktivitu takových lokalit pak významně zvyšovala také skutečnost, jestli
řekami procházely významné tahové cesty ryb. Tuto skutečnost dokládají četné
archeologické průzkumy prehistorických sídlišť v blízkosti řek, kde byly
mimo jiné nalezeny kosterní pozůstatky zejména lososů a jeseterů. Z těchto
archeologických nálezů můžeme číst, že pro prehistorického člověka měly
velký význam zejména diadromní ryby migrující mezi mořem a sladkou vodou a
to především z důvodu vysoké početnosti migrujících ryb, která byla s jinými
druhy nesrovnatelná. V tropických oblastech, kde jsou diadromní migrace
řídké, naproti tomu sehrály obdobnou roli druhy ryb s laterálními migracemi
v období záplav. Ryby, které zůstaly po opadnutí povodně v četných
sníženinách a příkopech zaplaveného území byly pro člověka snadnou kořistí.
Takový způsob lovu je stále praktikován mnoha původními kmeny tropické
Afriky, Jižní Ameriky a Asie.
Člověk v minulosti využíval říční ekosystémy více než ty ostatní. Je tedy
zřejmé, že dopady lidské činnosti se musely již v dávné minulosti v říčních
ekosystémech projevit. S postupných rozvojem schopností využívat přírodních
zdrojů docházelo také k prvním antropogenním změnám v krajině. Odlesňování a
stavba prvních přehrad a zavlažovacích systému působily v prostředí řek
změny, které měly přímý dopad také na migrující ryby. Nicméně rozsah těchto
změn byl do osmnáctého století příliš malý na to, aby významně ohrozil rybí
populace. Naopak obrovský rozvoj zemědělství a nástup průmyslové revoluce
stály u počátku drastických změn v říčních ekosystémech v posledních 300
letech. Důsledkem je mnohde zásadní narušení funkční integrity vodních toků,
čímž byly podmínky pro život mnoha druhů natolik znevýhodněny, že již
vyhynuly, nebo jim vyhynutí hrozí.
S postupným odstraněním mnohých tabu o přírodě v souvislosti s rozvojem vědy
a novými možnostmi člověka působit v krajině takovým způsobem, aby z toho
měl užitek, se změnil také pohled na řeky. Řeky přestaly být pouze
respektovaným živlem, zdroj obživy a transportu, ale staly se také místem
pro vypouštění odpadů, zdrojem vody pro průmysl a zejména rizikem v době
záplav, které bylo nutné eliminovat. Úpravy vodních toků mají dlouholetou
tradici a jejich cílem je právě omezit nekontrolované účinky vody, které
jsou často v rozporu s potenciálem využití území, které voda v době povodní
zasahuje. Koryta toků začala být napřimována, zkapacitňována, byla omezena
možnost rozliv vody do krajiny. Na tocích byly budovány rozsáhlé údolní
nádrže, které se měly stát jak zásobárnou vody, tak obřím rezervoárem v době
povodně. V současnosti je v ČR upraveno cca 30 % z celkové vodních toků,
která činí 76 000 km.
Důsledkem těchto vesměs jednostranných opatření bylo snížení morfologické
pestrosti koryt vodních toků. Tento faktor je důležitý zejména pro ryby s
výrazným teritoriálním chováním – například pstruh obecný. Nízké množství
úkrytů pak snižuje úspěšnost juvenilních jedinců pstruha při poproudových
migracích za účelem obsazení nových útočišť. V morfologicky pestrých tocích
jsou obecně migrace stanovištních druhů řídké z důvodu dostatečného prostoru
pro naplnění životních potřeb (např. parma říční). Naopak v uniformních
korytech se i tyto druhy musí vydávat za potravou z důvodů vysoké
konkurence, což zvyšuje riziko jejich ulovení predátorem a snižuje jejich
zdravotní kondici. Pro úpravy vodních toků je typická výstavba příčných
objektů často značné výšky, které jsou při protiproudových migracích pro
většin druhů nepřekonatelné. Účelem těchto objektů je snižovat podélný spád,
zajistit vzdutí vody pro jejich odběr a další. Významnou roli sehrávají tyto
objekty zejména v době třecích migrací. Tah ryb se u těchto objektu zastaví
a zpravidla pak dochází k jejich tření. Z důvodu ne vždy příznivých podmínek
pro tření a velkého množství jedinců ve tření je jeho úspěšnost významné
snižována a tím je ohroženo přežití populací. Diadromní ryby jako losos,
jeseter, úhoř tak v Evropě hluboko do vnitrozemí migrují jen na omezeném
množství lokalit (severní Evropa). Losos obecný k nám donedávna vůbec
nemigroval. Po zbudování rybích přechodů na četných překážkách na Labi a po
jeho umělém odchovu a vysazení do našich vod byl poprvé v roce 2002 opětovně
dospělý losos uloven v řece Kamenici. Přítomnost úhoře říčního v našich
vodách je v současnosti závislá výhradně na umělém vysazování. Možnosti
migrace úhořů jsou specifické a celá řada překážek na tocích je pro ně stále
neprostupná. Současně problematická je poproudová migrace úhoře z řek do
moří, neboť nepřekonatelnou překážkou pro ně mohou být přehrady. Navíc zde
hrozí riziko jejich poranění v turbínách vodních elektráren, které je s
ohledem na tvar jejich těla velice časté. Populace úhoře se tak právě v
důsledku významných změn omezujících realizaci jejich životního cyklu, stala
velmi ohroženou.
Fragmentace vodního toku příčnými překážkami má negativnívní vliv na
vnitrodruhovou genetickou diverzitu všech druhů ryb. Dochází k narušení
biologické kontinuity vodního prostředí, populace ryb oddělené neprostupnými
překážkami spolu nemohou komunikovat, sexuální kontakt se odehrává většinou
v rámci metapopulací ve fragmentovaných úsecích toku. Riziko příbuzenského
křížení je vysoké, dochází k postupné degeneraci genetické výbavy, čímž se
jedinci stávají náchylnější k onemocněním a schopnost přežití populací se
tím snižuje.
Obdobný negativní vliv na migrace ryb má znečištění vody a ovlivnění
hydrologického režimu. Silné trvalé znečištění toku představuje překážku,
která je pro migrující ryby nepřekonatelná. Stejně tak umělé snížení průtoku
vody k minimálním hodnotám, nebo až na nulový průtok neumožňuje migraci a
činí takto postižený úsek toku pro ryby neobyvatelným.
Ochrana migrujících ryb
Migrace ryby jsou zcela zásadním
biologickým projevem u většiny druhů a jejich narušením jsou ohroženy na
existenci mnohé druhy. Globální, ale i lokální neuvážené zásahy do vodního
prostředí způsobily změny v rybích společenstvech a poznání velkého rozsahu
těchto změn vedou ke snaze ryby i další živé formy účinnou formou chránit.
Nejdůležitějším nástrojem ochrany ichtofauny u nás je zákon č. 114/1992 Sb.
a prováděcí vyhláška č. 395/1992 Sb.. Podle těchto legislativních norem jsou
ryby chráněny jako volně žijící živočichové. Druhy, jejichž výskyt je v
určitém stupni ohrožení, jsou chráněni jako zvláště chránění živočichové a
to ve všech stádiích vývoje včetně jejich biotopu.
Ryby při svých migračních tazích překonávají často velmi dlouhé vzdálenosti
a jejich tahové cesty protínají hranice mnohá států ba i světadílů. Ochrana
migrujících populací pak vyžaduje mezinárodně koordinované aktivity, které
se stávají součástí mezinárodních smluv.
Literatura:
Lucas, M. & Baras, E. (2001): Migration of Freshwater Fishes. Blackwel
Publishing, Ames. 420 pp.
Baruš, V. & Oliva O. (1995): Mihulovci a ryby díl 1 a 2. Academia, Praha.
623 pp.
Lellák, J.& Kubíček, F. (1991): Hydrobiologie. Univerzita Karlova. Praha.
Hanel, L. & Lusk, S.(2005): Ryby a mihule české republiky. Rozšíření a
ochrana. ČSOP Vlašim.
Sigmund, L. a kol. (1992): Zoologie strunatců. Univerzita Karlova, Praha.
Begon M. a kol (1986): Ekologie. Jedinci, populace a společenstva.
Vydavatelství Univerzity palackého, Olomouc, 1997.
Moyle, P.B. & Cech, J.J. (1999): Fishes. An Introduction to Ichtyology.
Prentice Hall, New Persey.
Jungwirth, M.; Schmutz, S.; Weiss, S. (1999): Fish migration and Fish
Bypasses. Fishing News Books.

Obr. Výskok pstruha obecného při překonávání
překážky
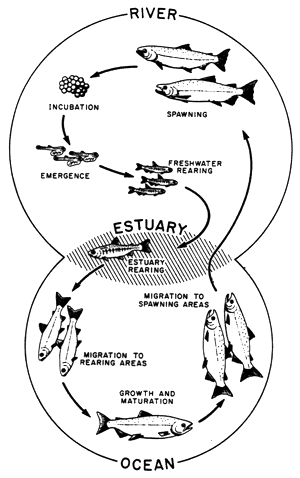
Obr. Schéma životního cyklu lososa obecného
Zdroj:
www.chemgapedia.de
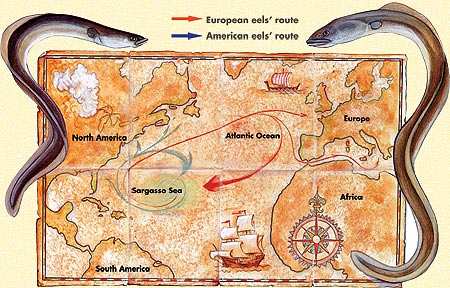
Obr.
Schéma
životního cyklu úhoře říčního
Zdroj:
www.harunyahya.com
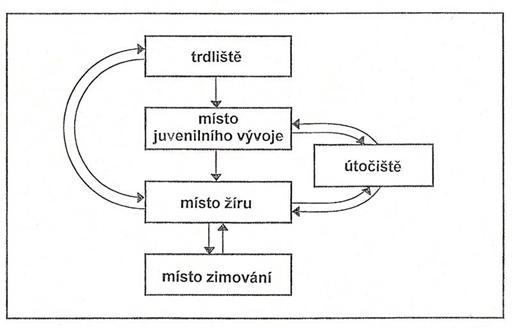
Obr. Schéma potamodromní
migrace

Obr.
Vranka
obecná
Zdroj: M. Štambergová

Obr.Řeka
Kamenice - od roku 2002 opět trdliště lososů o nás.
Zdroj: M.Štambergová

Obr.
Detail hlavy samce lososa
obecného uloveného v Kamenici,
Zdroj: M. Štambergová

Obr.
Tělo úhoře říčního, který
uhynul na následky zranění způsobeného turbínou vodní elektrárny
Zdroj:
www.glooskapandthefrog.org
|