|
Autotrofové Autotrofní organismus je takový, který dokáže přeměňovat anorganický uhlík na organický a jako zdroj energie obvykle využívá světlo (Smith, 1992). Perifyton, makrofyta a říční fytoplankton jsou třemi velmi rozdílnými skupinami autotrofních organismů, které se vyskytují v řekách a tocích. Perifyton, což je komplexní matrix autotrofů a heterotrofních mikroorganismů, která je přichycena na různých površích a substrátech, nacházíme prakticky ve všech vodních ekosystémech, ale zejména je častý na dně potoků a říček. Je důležitou součástí potravy pro bezobratlé a některé ryby, a může také fungovat jako důležitý sorbant kontaminujících látek.
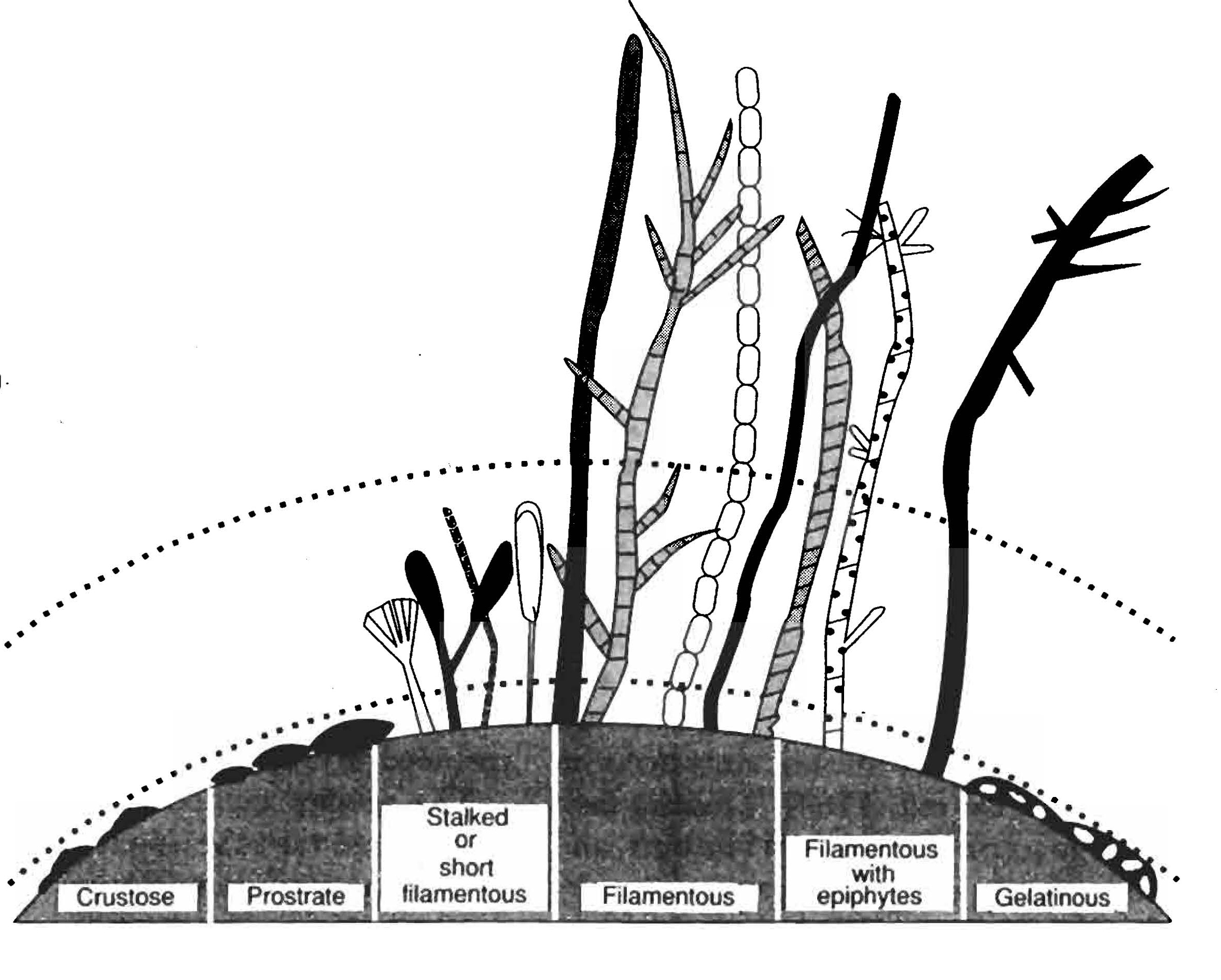
Obr. Hlavní růstové formy perifytonu ilustrují značnou variabilitu jejich tvarů a vertikálního rozvrstvení. Tyto strukturální vlastnosti podle očekávání určují jejich náchylnost ke spásání konzumenty. Zdroj: Allan, 1995 Perifyton se skládá z množství rozsivek, zelených řas, sinic a několika dalších skupin organismů, a nachází se prakticky na každém podkladu v tekoucí vodě, včetně kamenů (epilitické formy), měkkých sedimentů dna (epipelické formy, bentos) a makrofyt (epifytické formy). Naprostá většina druhů perifytonu patří mezi rozsivky, běžné jsou však také zelené řasy a sinice, které mohou za určitých podmínek i dominovat. Druhové složení perifytonu se v tocích mírného pásma mění poměrně pravidelně podle ročního období. Během zimy, jara a časného léta typicky dominují rozsivky. Složení jejich druhů se během tohoto období mění a jejich největší celkové množství bývá většinou na jaře, kdy je intenzita světla už dostatečně vysoká a ještě nejsou vyvinuty listy stínící pobřežní vegetace. Populace zelených řas a sinic narůstají během léta, ale to celková biomasa bentických autotrofů klesá kvůli zvětšujícímu se zastínění, a krátce stoupne zase až na podzim (Moore). Sinice obvykle rostou jako epilitické a epipelické, mezi epifyty převládají spíše rozsivky a zelené řasy. Množství perifytonu se mění jak sezónně, tak prostorově, a to díky mnoha faktorům, z nichž každý může být za určitých podmínek ten nejpodstatnější a převládající. Vysoký průtok často omezuje růst perifytonu na klidnější období tím, že strhává organismy, převrací kameny a obrušuje povrchy. Světlo může být limitujícím faktorem zejména v potocích pod hustými korunami stromů v lese. Stále více se ukazuje, že velmi běžná je limitace živinami, nejčastěji nedostatkem fosforu. Malí autotrofové jsou také často konzumováni herbivory a spásání pro ně může znamenat značné ztráty produkce. Makrofyta, kam patří některé velké řasy, mechy a cévnaté rostliny, se nacházejí v tekoucích vodách obvykle jen tam, kde není ani velká hloubka, ani proud. Středně veliké řeky, kanály a okraje řek dokáží podporovat jejich největší biomasu. Rozlišujeme čtyři hlavní skupiny makrofyt podle růstové formy: emerzní formy v blízkosti břehů a na mělčinách, druhy s plovoucími listy na okrajích pomalu tekoucích řek, volně plovoucí rostliny, jaké například nacházíme ve velkých shlucích ve velkých tropických řekách, a submerzní druhy, které se přichycují k podkladu. Délka období růstu, množství světla a rychlost proudu jsou pro vodní makrofyta hlavními limitujícími faktory, jak je identifikuje Westlake (1975). Zastínění okolní vegetací snižuje osvětlení hladiny toku o 35-95% a může způsobit naprosté vymizení mechorostů. Zakalení vody, v kombinaci se zastíněním a druhově specifickými požadavky na množství světla, mohou inhibovat růst makrofyt v hlubších úsecích řeky. Úspěšné přežívání populací makrofyt v silném proudu je umožněno několika adaptacemi. Většinu druhů lotických makrofyt charakterizují pevné a pružné stonky a listy, přichycení se přídatnými kořeny, oddenky a šlahouny, a vegetativní rozmnožování (Hynes, 1970; Westlake, 1975). Spásání živých rostlin má ve většině případů jen malý vliv, a většina produkce se po odumření rostlin dostává mezi detrit. Kromě mechorostů většina vodních makrofyt v zimě ve vodním sloupci odumírá. Fytoplankton, zahrnující buňky a kolonie malých autotrofů ve vodním sloupci, tvoří populace schopné samostatné existence jen za určitých podmínek, které se obvykle nacházejí ve velkých nížinných tocích. První z nich je aby čas, po který zůstává plankton na určitém místě toku, byl dostatečně dlouhý na to, aby produkce biomasy byla větší než její odnos proudem. Živiny obvykle netvoří limitující faktor říčního planktonu, kterým však často bývá světlo, a to všude tam, kde je proud dostatečně pomalý, aby umožňoval bohatý rozkvět fytoplanktonu. Hloubka, do které světlo proniká, je obvykle mnohem menší než hloubka těchto velkých kalných řek, a protože je vodní sloupec obvykle velmi dobře promíchán, nachází se fytoplankton po většinu času v podmínkách nedostatečného nebo dokonce nulového osvětlení. V reálné situaci může být čas vhodný pro fotosyntézu fytoplanktonu tak krátký, že by jeho populace v toku nemohly přežít bez přispění mělkých oblastí, zátok nebo záplavových jezírek. Ve srovnání se stojatou vodou, obsahující přibližně stejné množství živin, je biomasa říčního fytoplanktonu vždy podstatně menší. Také se zdá, přestože naše znalosti jsou v této oblasti skrovné, že ani spásání fytoplanktonu není silným limitujícím faktorem. Na rozdíl od stojatých vod, kde je fytoplankton obvykle limitován určitou kombinací omezených zásob živin a spásání, jsou tedy v řekách tyto faktory mnohem méně důležité. Současné poznatky naznačují, že dominujícím osudem produkce fytoplanktonu je spíše odnos proudem, než její přeměna na místě vzniku.
Obr. Schéma srovnávající vliv hloubky promíchávání na primární produkci v jezerech a v řekách. V jezerech (a) se vytvoří teplotní bariéra mezi povrchovou a hlubokou vodou, která omezuje promíchávání na několik metrů. V řekách (b) teplotní stratifikace kvůli víření proudu nemůže vzniknout a vodní sloupec se promíchává až ke dnu. Transparentnost, získaná měřením pomocí Secchiho disku jako hrubé měření průniku světla použitelného pro fotosyntézu, dosahuje v mezotrofických a eutrofických jezerech 1-8m, a může být mnohem větší v čistých, málo produktivních jezerech. Velké řeky jsou běžně hluboké 5-20m, ale kvůli vysokému obsahu částic sedimentů je pronikání světla obvykle omezeno na svrchní 1-2m. Zdroj, Allan, 1995 Relativní důležitost těchto tří velmi odlišných skupin primárních producentů se značně mění s měnící se velikostí řeky a s podmínkami v ní. U všech tří skupin můžeme zjišťovat limitující faktory a také velikost a další osud jejich produkce. |