|
Trofické vztahy v jezeře Na rozdíl od řek je v jezerech většina chemické energie pro organismy fixována pomocí fotosyntézy. Světla s hloubkou rychle ubývá a když poklesne pod 1% povrchového osvětlení, fotosyntéza není možná. Při této intenzitě osvětlení převládne respirace nad fotosyntézou, buňky řas dále nerostou a bez jejich růstu se nevytvářejí zdroje potravy pro konzumenty. Proto tato intenzita osvětlení určuje hranici mezi eufotickou (fotickou) a afotickou zónou (kompenzační hladina fotosyntézy) (Brönmark and Hansson 2002). Hloubka eufotické zóny určuje, jestli jsou buňky fytoplanktonu vystaveny větší či menší intenzitě osvětlení, jaká je konečná distribuce primární produkce a jaká je velikost biomasy fytoplanktonu. Pohyby vody v epilimniu, způsobené vířením nebo vlnami, způsobují cirkulaci fytoplanktonu v této vrstvě. Když v jezeře neexistuje teplotní stratifikace (epilimnion), buňky řas mohou klesnout do příliš velké hloubky, kde pro jejich růst není dostatek světla (Kalf 2002). Pokud by tedy v hlubokých jezerech tato stratifikace nebyla, dostával by se fytoplankton (jehož buňky jsou příliš malé na to, aby kontrolovaly svoji pozici ve vodním sloupci a jsou jen pasivně unášeny pohybem vody) do hloubek hodně pod eufotickou zónou. Ve volné vodě jezer (pelagiálu) by tak byla příliš nízká fotosyntéza, než aby to umožňovalo existenci života. Růst planktonních řas by byl omezen na ty oblasti jezer, kde je hloubka dna menší než eufotická zóna – podobně jako tomu je u makrofyt a přisedlých řas. Stratifikace umožňuje růst fytoplanktonu v pelagiálu hlubokých jezer, ale zároveň také způsobuje odebírání živin z epilimnia. Jejich nedostatek může následně způsobit omezení růstu fytoplanktonu. Za běžných podmínek jsou vody pelagiálu vždy méně produktivní než vody litorálu, kde živiny také mohou klesnout na dno, ale z něj jsou bentickými organismy znovu vráceny do vodního sloupce.
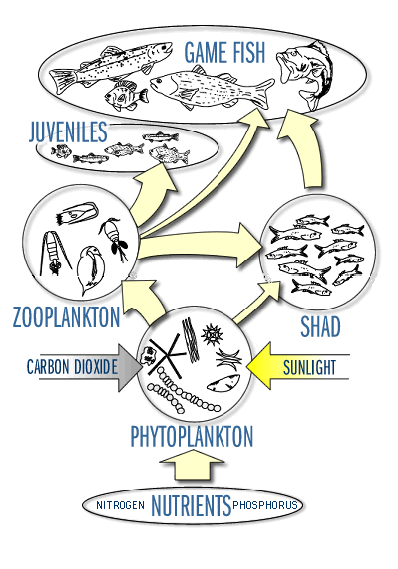
Obr. Biologická společenstva v jezerech mohou být organizována podle konceptu potravních řetězců a sítí, což nám umožňuje porozumět fungování ekosystému. Ve většině jezer můžeme rozlišit tři různé potravní sítě – litorální, planktonní a mikrobiální (Brönmark and Hansson 2002). Některé druhy organismů, zejména pohyblivé ryby, se však mohou účastnit více typů řetězců a tak integrovat tok energie v jezeře.
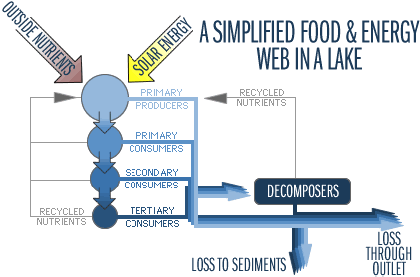
Obr. Potravní sítě můžeme popisovat buď jako toky energie nebo živin (uhlíku, dusíku nebo fosforu) V litorálu je mnoho primárních producentů, ať už se jedná o makrofyta („velké“ rostliny), fytobentos (buňky bentických řas jsou také obvykle větší než fytoplanktonní) nebo perifyton. Tyto litorální zdroje potravy mohou být předány do vyšších trofických úrovní skrze velké bezobratlé, kteří se bentickými rostlinami živí. Tento trofický řetězec může být velmi produktivní a efektivní, ale ve velkých jezerech je omezený na relativně malý prostor. Pelagičtí primární producenti (fytoplankton) jsou malí. Jejich velikost vyžaduje využití malými konzumenty – zooplanktonem. Nicméně i to může být základ důležitého potravního řetězce, mířícího do vyšších trofických úrovní, zejména jsou-li přítomny zooplanktonožravé ryby. V každém případě je zooplankton důležitý pro potěr a malé ryby, které nemohou pozřít větší kořist. Dekompozitoři, zahrnující bakterie, houby a jiné mikroorganismy, jsou důležitou součástí mikrobiální potravních sítí. Živí se zbytky vodních organismů a tím, že rozkládají organickou hmotu, ji navracejí do anorganické formy. Část těchto rozložených látek je následně znovu využívána jako živiny, jako třeba fosfor ve formě fosforečnanů (PO43-) a dusík ve formě amoniaku (NH4+), které jsou snadno dostupné pro nový růst rostlin. Uhlík se obvykle uvolňuje jako oxid uhličitý (CO2), který snižuje pH vody v blízkosti dna. V anoxických podmínkách (bez přítomnosti kyslíku) je někdy uhlík vylučován jako plynný metan (CH4). Dekompozitoři mohou být nalezeni ve všech biologických zónách jezer, avšak nejčastěji převládají v hypolimniu, kde je hojnost mrtvé organické hmoty (z odpadních vod, mrtvého planktonu, odumřelých makrofyt a organických sedimentů, spláchnutých z povodí). Oxidace této hmoty dekompozitory (respirace) v hypolimniu způsobuje v průběhu léta vyčerpání rozpuštěného kyslíku, což může vést k anoxickému stavu. V hypolimniu není žádný zdroj kyslíku, který by nahradil jeho ztráty při dekompozici: stratifikace zabraňuje atmosférickému kyslíku, aby pronikl pod termoklinu, a nedostatek světla obvykle znemožňuje fotosyntézu. Výsledná anoxie má závažný vliv na chemické i biologické procesy v jezerech. Mikrobiální potravní řetězce využívají DOM a rozpuštěné živiny a udržují je v oběhu. Nejsou tak efektivní jako ostatní, ale pro koloběh živin jsou velmi důležité. Obr. Model koloběhu živin a toku energie v ekosystému jezera. Meteorologické (hydrologické), geologické a biologické vstupy se dostávají do lenitického systému z jeho povodí. Živiny a energie, které do systému vstupují, nebo jsou v něm vyprodukovány, se pohybují po mnoha drahách. Část energie a živin je fixována do sedimentů dna. Zdroj : Smith, 1992 |